Clase 2. Tansportadores (Carriers).
|
|
|
![]()
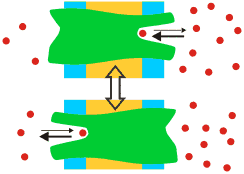
Son moléculas que llevan a cabo procesos de transporte empleando como fuente de energía el potencial electroquímico de al menos uno de los solutos transportados, y mediante un cambio conformacional del propio transportador, tras la formación de un complejo reversible transportador-soluto.
|
|
El sitio de unión del soluto se encuentra alternativamente a un lado y a otro de la membrana. Este es un ejemplo de Uniporte |
|
En realidad, el complejo transportador-soluto(s) es simplemente un caso particular de la formación de complejos proteína-ligando(s), por lo que son de aplicación todos los conceptos estudiados en el tema correspondiente, particularmente la existencia de un flujo máximo de soluto (análogo a la velocidad máxima de una reacción enzimática), cuando todas las moléculas de transportador tienen ocupado su sitio de unión. Muestran saturación respecto a la concentración de soluto. Del mismo modo que en el caso general proteína-ligando, se puede definir una K50 que corresponde a la concentración de soluto que permite un flujo mitad del máximo.
|
|
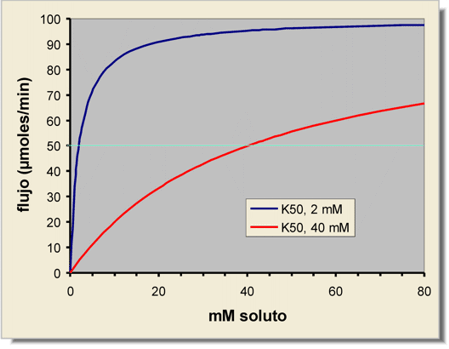
| Ejemplo teórico de flujo (expresado como moles por unidad de tiempo) a través de una membrana, en función de la concentración de soluto en el compartimento de salida. La concentración en el de llegada se supone 0 (flujo inicial). Se representan los resultados para dos solutos diferentes, que se diferencian en sus afinidades hacia el transportador. En ambos casos, el flujo máximo, que depende de la velocidad a la que el transportador cambia de conformación, se supone igual a 100 µmoles soluto transportado/minuto |
La velocidad específica de transporte es de uno a dos órdenes de magnitud menor que para los pocos y canales, ya que en este caso el transportador tiene que mover físicamente al soluto de un lado a otro de la membrana, mientras que en los poros las moléculas se mueven directamente por difusión.
Hay dos tipos fundamentalmente diferentes desde el punto de vista bioenergético:
Uniporte: Se transporta un
único soluto, a favor siempre de su potencial electroquímico. Se trata de un
caso de Difusión Facilitada
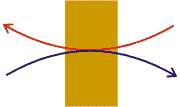
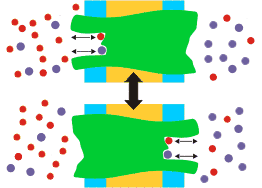
Cotransporte (o Transporte activo secundario): Transporte mecanísticamente acoplado de dos (o más raramente tres) solutos. Al menos uno de los solutos se transporta a favor de su potencial electroquímico, lo que suministra la energía necesaria para transportar al segundo soluto en contra de su potencial electroquímico. Si se transportan ambos solutos en el mismo sentido se trata de Sinporte; si se transportan en sentidos opuestos, Antiporte.
|
|
Antiporte: Los dos solutos se transportan en direcciones opuestas |
|
|
|
|
|
|
|
Sinporte: Los dos solutos se transportan en la misma dirección |
|
|
En estos esquemas se supone que el transporte del soluto "rojo" es exergónico, mientras que el transporte del soluto "azul" es endergónico |
||
Es importante destacar el concepto de acoplamiento mecanístico. Evidentemente, el soluto que consume energía libre no se puede transportar en ausencia del soluto que la proporciona, pero en los cotrasportadores el soluto cuyo transporte está favorecido termodinámicamente NO se transporta en ausencia del otro; en este caso, no por falta de energía, sino porque el transportador no funciona. Esta restricción evita que se disipe inútilmente el potencial electroquímico de este soluto.
Se denominan procesos de Transporte activo secundario porque:
a) Uno de los solutos se transporta en contra de su potencial electroquímico, y
b) Se requiere un proceso de transporte activo primario para que el potencial electroquímico del primer soluto sea suficientemente elevado para permitir el transporte del segundo.
Los procesos de cotransporte son, con mucho, más frecuentes que los de uniporte. De hecho, el uniporte parece ser un proceso excepcional, limitado al transporte de azúcares en vertebrados, levaduras y alguna bacteria. Todos los transportadores pertenecen a la subclase 2.A, de la que se han descrito un total de 81 familias. La subclase 2.B corresponde a transportadores no sintetizados en ribosomas, de los que la valinomicina es un conocido ejemplo.
Ejemplos de transportadores:
El grupo más importante es 2.A.1, denominado la Principal Superfamilia de facilitadores (MFS, Main Facilitator Superfamily), con alrededor de mil miembros secuenciados. Se encuentran en todos los grupos de seres vivos. La mayor parte de ellos están formados por un haz transmembrana de 12 o 14 hélices a. En esta superfamilia existen diversas familias. Algunos ejemplos de estos transportadores son los siguientes:
Uniporte de glucosa en animales, para el que hay varias proteínas diferentes: GLUT1 (2.A.1.1.28), general, del que existe un modelo teórico de su estructura terciaria ; GLUT2, (2.A.1.1.29); GLUT3, (2.A.1.1.12), que se encuentra en cerebro, y GLUT4. Se diferencian en los tejidos donde se expresan, en su afinidad hacia la glucosa y en el requerimiento o no de insulina para su presencia en la membrana citoplásmica:
![]()
|
|
|
La concentración "normal" de glucosa en la sangre es del orden de 5 mM (línea vertical magenta). El flujo de glucosa dependiente de GLUT1 es prácticamente constante en el rango normal de concentraciones de glucosa en sangre (en blanco en la figura), pero cae rápidamente a concentraciones de glucosa inferiores a 3 mM. Por su parte, el flujo dependiente de GLUT2, debido al elevado valor de su K50, varía linealmente con la concentración de glucosa. En el hígado, la glucosa entra (o sale) en función directa de sus concentraciones intrahepática y sanguínea. Por último, GLUT4 incrementa su presencia en las membranas de los adipocitos y del músculo en presencia de insulina (flecha en la figura): la insulina aumenta 20 veces la actividad de GLUT4 en la membrana de adipocitos. En ausencia de la hormona el transporte de glucosa en estos tejidos es bajo. |
En levaduras y en la bacteria Zymomonas hay también sistemas de uniporte para azúcares.
2.A.1.2 : Sistemas de antiporte droga-H+, por ejemplo antiporte tetraciclina-H+ en bacterias (y otros sistemas similares específicos para diferentes antibióticos). Muy importantes como mecanismo de resistencia a antibióticos.
Otros ejemplos son los diferentes sistemas de sinporte aminoácidos-Na presentes en animales; o el sistema de sinporte para múltiples vitaminas dependiente de sodio, también de animales, etc. De hecho, la mayor parte de los sistemas de captación de nutrientes del exterior celular o de exportación de compuestos de desecho en eucariotas son sistemas de cotransporte. También son muy frecuentes en procariotas. En animales el soluto que dona la energía para el transporte desde el exterior al interior celular es el sodio; en plantas, hongos y en bacterias (excepto en las alcalófilas) este papel lo cumple el H+.
|
|
|