Concentración constante de ligando
![]()
Consideraremos que la concentración de ligando es constante, y estudiaremos el comportamiento individual de una molécula de proteína. Recuerde que cada molécula de proteína une o libera el ligando independientemente de las demás; la vida media de cada complejo es el inverso de la Kd, pero ese es un valor promedio; a veces el complejo se disociará mucho antes, y otras veces mucho después de esa vida media. Por otra parte, a lo largo del tiempo, la saturación media de una molécula de proteína equivale a la fracción de saturación global de toda la población definida por la ecuación:
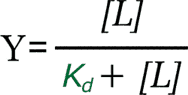
es decir, si la fracción de saturación es del 0,25 (25%) en el conjunto de la proteína, dado un tiempo suficientemente largo UNA molécula de proteína estará saturada el 25% de dicho tiempo.
Supongamos un receptor hormonal que es una proteína de membrana y que como consecuencia de la unión de la hormona sufre un cambio conformacional que resulta básico para la transmisión del mensaje hormonal al interior de la célula. Este caso es muy corriente, y de hecho, excepto las hormonas esteroideas, la mayor parte de las hormonas actúan por mecanismos de este tipo, mediante un "segundo mensajero" intracelular. Veremos ejemplos con mas detalle más adelante, pero un caso típico es el del control por la adrenalina del metabolismo del glucógeno, en el que el segundo mensajero es el AMP cíclico (AMPc):
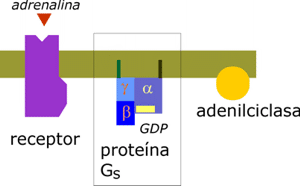 |
Consideremos este caso, donde la hormona es un soluto de bajo peso molecular (211). Supongamos que existen dos posibles tipos de receptores: Tipo A, cuya Kd para la adrenalina es 1 mM, y Tipo B, cuya Kd es 1 mM. Cada uno de estos receptores puede encontrase en dos estados, 1, sin adrenalina unida y 2, con adrenalina unida y con la capacidad de transmitir la señal al interior celular. Es importante tener en cuenta que la transmisión de la señal requiere algún tipo de proceso (en este ejemplo, la interacción con las proteínas Gs y el cambio de conformación de las mismas), proceso que puede ser rápido, pero no es instantáneo. Dado que la adrenalina es un soluto pequeño podemos asumir que el valor de K1 es de 108 M-1.s-1. Veamos como se comportan ambos tipos de receptores; para simplificar un poco más asumamos un 50% de saturación.
| Receptor | Tipo A | Tipo B |
| Kd | 1 mM | 1 mM |
| Vida media del complejo receptor-hormona | 0,01 s | 0,00001 s |
| gramos de hormona necesarios para conseguir una saturación del 50%, asumiendo un volumen sanguíneo de 5 litros | 1,05 mg | 1,05 g |
Aunque si la concentración de adrenalina tiene el valor adecuado los receptores alcanzan el 50% de saturación en cada caso, no olvidemos que se trata de un valor promedio y que la realidad es que los receptores están uniendo y liberando continuamente el ligando. Para los receptores del tipo A, la duración promedio del complejo para cada receptor individual es suficientemente grande para que cambie de conformación y se una y active la proteína Gs, que se mueve al azar por la membrana. En resumen, en 10 milésimas de segundo hay tiempo para que sucedan todos estos acontecimientos, e incluso para que se repitan varios ciclos de activación de la proteína Gs. Sin embargo, para los receptores del tipo B, aunque la saturación promedio sea exactamente la misma, cada receptor está unido en promedio 10 millonésimas de segundo a la hormona, mil veces menos tiempo que para los receptores del Tipo A. ¿Es tiempo suficiente?. Para el cambio conformacional, desde luego. Pero es evidente que los siguientes pasos del proceso son menos probables dado que puede suceder que no dé tiempo a que una proteína Gs interactúe con el receptor durante el breve plazo en que se encuentra en la conformación apropiada, por lo que cabe esperar que muchos de los acontecimientos de unión hormona-receptor no se traduzcan en activaciones de la proteína Gs.
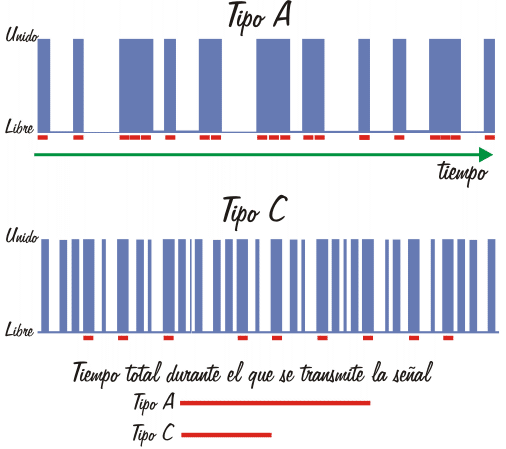 |
Esquema gráfico de la discusión anterior. En este ejemplo, la vida media del complejo receptor de tipo C-ligando es aproximadamente un tercio de la del receptor de tipo A-ligando. El tiempo necesario para que se transmita un ciclo de la señal se representa como un rectángulo rojo, y es constante. En ambos casos el tiempo que el receptor permanece unido al ligando es un 50% del total. Evidentemente, cuanto mayor es la vida media del complejo mayor es la probabilidad de que se transmita la señal. |
En resumen, el proceso es menos eficiente cuanto menor es K-1 (o mayor Kd, si la constante de asociación K1 es similar). No sólo porque hace falta mucha mas cantidad de la hormona para alcanzar el 50% de saturación de los receptores de la célula, sino porque además, una vez formado el complejo, es menos probable que ese complejo tenga tiempo de transmitir la señal al interior celular. No es pues de extrañar que los valores reales de Kd para los receptores adrenérgicos en humanos estén en el rango de 0,25 a 4 mM.
La importancia del tiempo que se mantiene unido el ligando a la proteína será, evidentemente, más significativa si la transmisión de la señal es lenta respecto a la unión del ligando. Si es muy rápida (por ejemplo la señal consiste en entrada de iones a través de un canal, que se abre o cierra según tenga o no un ligando unido), este efecto es probablemente irrelevante.
![]()