![]()
|
|
|
Aunque la formación del complejo P.L. se define habitualmente empleando el valor de Kd, merece la pena comentar brevemente el significado físico y los límites de variación de las constantes individuales K1 y K-1.
K1 es la constante de velocidad de segundo orden para la unión de la proteína al ligando:
![]()
Esta constante no puede superar un valor máximo, que viene dado por la velocidad a la que se pueden encontrar ambas moléculas cuando se desplazan al azar en disolución, y que depende de sus respectivos Coeficientes de Difusión . En un medio líquido homogéneo, este valor máximo se encuentra en el rango 108 a 109 M-1.s-1. En la mayor parte de los casos los valores de K1 van a ser inferiores a este máximo, ya que no todos los choques posibles entre dos moléculas van a ocurrir con la orientación adecuada para que se forme el complejo. De hecho, en la mayor parte de los casos los valores de K1 se encuentran en el rango 105 - 108 M-1.s-1. Por otra parte, cualquier factor que disminuya el valor de los coeficientes de difusión (aumento de la viscosidad del medio, aumento del tamaño -por agregación, por ejemplo- de los reactantes), va a traducirse en una disminución del valor de esta constante de velocidad. Como ejemplo, el valor de K1 para la asociación proteína-proteína entre la elastasa y la a1-antitripsina se ha estimado en 6,7.107 M-1.s-1.
K-1 es la constante de velocidad de primer orden para la separación del complejo P.L.
![]()
En este caso, y como ocurre para cualquier proceso cinético de primer orden, hay una relación sencilla entre la vida media del complejo y el valor de K-1. NO se debe confundir con el periodo de semidisociación (análogo al periodo de semidesintegración, o semiperiodo, en la desintegración radiactiva).
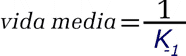 |
Tiempo promedio de existencia de un complejo P.L. | |
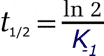 |
Tiempo requerido para que la concentración inicial del complejo P.L se reduzca a la mitad, suponiendo que no se sintetice de nuevo. |
Hasta los últimos años la determinación de los valores individuales de estas constantes era un proceso bastante complicado, por lo que la mayor parte de los datos existentes se refieren exclusivamente a valores de Kd. Sin embargo, la determinación directa de las constante K1 y K-1 se puede llevar a cabo por técnicas especiales, como la resonancia de plasmones, y cada vez es mayor el número de equilibrios que se están estudiando a este nivel.
Kd y duración del complejo PL
Dado que Kd, K-1 y K1 están relacionadas, se puede calcular la vida media de existencia de un complejo P.L. si conocemos Kd y la constante de asociación K1. Por ejemplo, para el caso del par elastasa.a1-antitripsina, cuya Kd tiene un valor aproximado de 10-11 M, y teniendo en cuenta el valor de K1 anteriormente expuesto, se puede calcular que el valor de K-1 es de 6,7.10-4 s-1, lo que corresponde a una vida media del complejo de unos 1500 segundos.
Puede ser curioso calcular los valores de vida media que corresponden a diferentes valores de Kd. Evidentemente, necesitamos conocer los valores reales de K1 para cada caso concreto, pero aunque sea un cálculo poco exacto los resultados no dejan de ser interesantes:
| Tabla 1 |
 |
| Vidas medias (inversos de K-1) para varios valores de Kd, asumiendo dos posibles valores de K1. |
En la tabla 1 se puede apreciar cómo la disminución en el valor de la constante de disociación del complejo se relaciona con un aumento del tiempo de vida medio del mismo, que pasa del rango de micro o milisegundos a periodos de tiempo mucho más prolongados. Esto, además de como curiosidad, puede tener implicaciones funcionales. Por ejemplo, unos complejos P.L de gran importancia son los formados por una hormona unida a su receptor; el valor de Kd puede dar una idea aproximada del tiempo durante el cual el receptor se encuentra activado al estar unido a la hormona correspondiente. Un valor bajo de Kd no supone sólo que el receptor requiere menor concentración de hormona para saturarse sino que, a igualdad de otros factores, se encuentra activado durante más tiempo, tanto si la concentración de ligando es constante (ejemplo 1) como si la misma disminuye rápidamente (ejemplo 2). Un ejemplo reale sobre la importancia del tiempo en la asociación de ligandos se presenta en el ejemplo 3.
|
|
|