PROCESOS QUIMIOSMÓTICOS.
|
|
![]()
Cuando una membrana delimita un compartimento
cerrado, y se establece por algún mecanismo una diferencia de concentración de
un soluto o de potencial, si es el caso- a ambos lados de la membrana, el
transporte del soluto a favor de su potencial es exergónico, esto es, libera
energía libre. Esta es una situación de alta energía de transporte análoga a
la situación de alta energía de hidrólisis que tienen los enlaces pirofosfato
que se encuentran en el ATP. En ambos casos cuando el sistema (gradiente de
soluto; ATP) cambia de estado (el soluto se transporta; el ATP se hidroliza) se
libera energía que, de existir un mecanismo de acoplamiento adecuado, se puede
aprovechar. Esta similitud está en la base del modelo quimiosmótico de Peter
Mitchell, que fue quien por primera vez desarrolló la idea de la posible
transducción de energía entre gradientes de soluto el componente
"osmótico"- y
reacciones químicas.
De hecho, durante mucho tiempo se consideró al ATP la moneda energética de la célula, que transfería energía libre de unas reacciones a otras. Pues bien, las células (todas) tienen dos tipos de moneda energética, dos tipos de mecanismos de almacenamiento inmediato y de transducción de energía: el sistema ATP-ADP y los gradientes de solutos. Podemos extender esta metáfora, diciendo como V. Skulachev que las células pueden usar efectivo el ATP- y cheques: los gradientes de solutos. Y ambos tipos de "dinero" son intercambiables entre sí. Por ejemplo, el gradiente de H+ a través de la membrana plasmática de bacterias se puede crear mediante hidrólisis de ATP o mediante reacciones redox, y se puede emplear para sintetizar ATP, para transportar solutos en contra de su potencial electroquímico o para desplazar una célula bacteriana mediante rotación de su flagelo. Hay una libre interconversión entre energía química, de transporte o quimiosmótica, y mecánica.
Por otra parte, la dependencia del transporte activo secundario de la creación de gradientes de solutos mediante el transporte activo primario da lugar a la creación de circuitos de entrada y salida de solutos, que pueden ser bastante complicados. A continuación veremos algunos ejemplos.
![]()
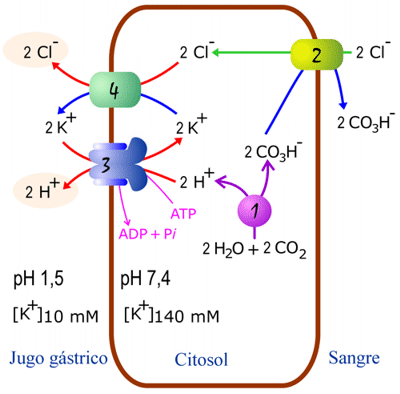
Acidificación del jugo gástrico
El jugo gástrico de los vertebrados tiene un valor de pH extremadamente bajo; en mamíferos, se encuentra entre 1 y 2. Esta elevada acidez se debe a la excreción de HCl por parte de células existentes en la mucosa gástrica. La exportación de HCl por estas células es un bonito ejemplo de integración de diversos sistemas de transporte.
El ión Cl-`proviene de la sangre; el H+ proviene del agua; ambos se deben exportar al jugo gástrico en contra de sus potenciales electroquímicos. El proceso se muestra a continuación:
|
|
|
|
1: Anhidrasa carbónica 2: Antiporte electroneutro cloruro-bicarbonato 3: H+K+ ATPasa electroneutra. 4: Sinporte electroneutro cloruro-potasio En rojo transporte endergónico; en azul, transporte exergónico
|
|
![]()
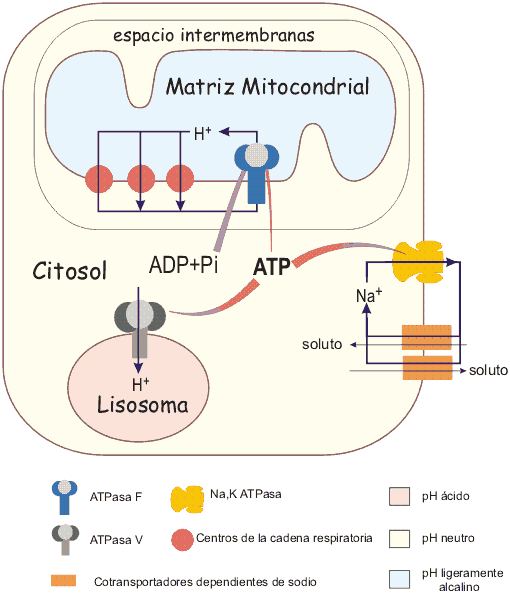
Circuitos quimiosmóticos en una célula animal
En una célula animal típica, la mayor parte del ATP se sintetiza a partir de ADP y fosfato mediante la fosforilación oxidativa mitocondrial. En este proceso los tres centros redox de la cadena respiratoria mitocondrial bombean hidrogeniones desde la matriz mitocondrial al espacio intermembranas, desarrollando una fuerza protonmotriz del orden de - 180 mV. Esa fuerza protonmotriz permite la síntesis de ATP mediante la ATPsintetasa F. El ATP se puede emplear, entre otras muchas reacciones, para establecer los gradientes de sodio y potasio a través de la membrana plasmática de la célula (mediante la bomba de sodio-potasio del tipo P), y para acidificar los lisosomas mediante una ATPasa V. El gradiente de sodio se emplea como fuente de energía para el transporte de solutos a través de la membrana plasmática mediante diferentes procesos de cotransporte. Obviamente esto es un modelo muy simplificado: hay otras ATPasas de tipo P y de tipo ABC en los diferentes sistemas de membrana.
![]()
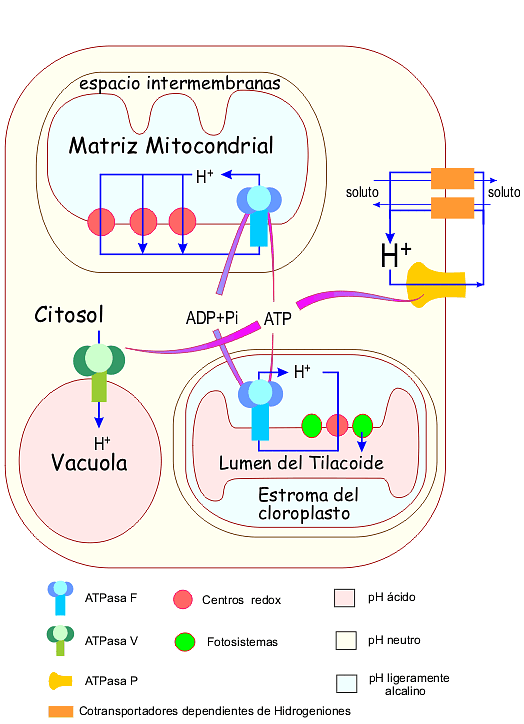
Circuitos quimiosmóticos en una célula vegetal
En una célula vegetal NO fontosintética la situación es similar a la de la célula animal (el ATP se obtiene fundamentalmente mediante la fosforilación oxidativa), con la diferencia de que el soluto que se utiliza para el cotransporte a través de la membrana plasmática es el ión H+, en vez del Na+. El gradiente de H+ en la membrana plasmática se crea mediante una ATPasa de tipo P. En hongos y levaduras la situación es similar, excepto cuando están en anaerobiosis. En ese caso, el ATP se produce exclusivamente por la fosforilación a nivel de sustrato que ocurre en la fermentación. En una célula fotosintética existe un tercer compartimento, el cloroplasto, que sintetiza ATP. En este caso se crea un gradiente de pH a través de la membrana del tilacoide, entre el estroma del cloroplasto y el lumen del tilacoide. El ATP se puede emplear en los procesos biosíntéticos del cloroplasto, o ser exportado al citosol.
![]()
FIN DEL TEMA